 Anatomi i morfologia Anatomi i morfologia
W tym dziale znajdziesz omówienie następujacych elementów budowy spodoustych Elasmobranchii:
Szkielet:
Tkanka chrzęstna
Płetwy
Kręgosłup i żebra
Czaszka chondrocranium
Łuski plakoidalne
Anatomia miękka:
Układ moczowo-płciowy
Układ pokarmowy
Układ krwionośny i serce
Układ nerwowy
Układ pokarmowy
Większość elementów składowych układu pokarmowego u spodoustych - Elasmobranchii składa się z tych samych elementów co u innych kręgowców tj. z otworu gębowego, przełyku, żołądka oraz jelita.
Pierwszym odcinkiem układu pokarmowego jest otwór gębowy. Znajdują się tu zęby U rekinów brak jest języka w paszczy. Przełyk ma dużą średnicę i grubą ścianę. Na wewnętrznej ścianie przełyku są liczne kosmki. Żołądek u spodoustych jest jednokomorowy. Ściana żołądka posiada liczne fałdy zwiększające jego powierzchnię. W jelicie znajduje się zastawka spiralna, która zwiększa powierzchnie chłonną jelita. W pobliżu końcowego odcinka przewodu pokarmowego mają ujście gruczoły solne wydalające solankę z wody morskiej.
Zakończeniem układu pokarmowego u wszystkich chrzęstnoszkieletowców jest kloaka czyli wspólne ujście układu pokarmowego, rozrodczego i wydalniczego (podobny twór występuje u stekowców, gadów i ptaków).
Wątroba u spodoustych jest monstrualnych rozmiarów, trójpłatowa. W wątrobie mogą być gromadzone duże ilości tłuszczów - diacylogliceroli. Związane jest to z pełnieniem funkcji narządu hydrostatycznego przez ten organ (u spodoustych brak jest pęcherza pławnego). Między płatami wątroby znajduje się mały woreczek żółciowy. W okolicach przedniego odcinka jelita zlokalizowana jest trzustka.
Typy zastawki jelita występujące u spodoustych (wg White 1937):
- typ spiralny - jest to typowa zastawka (Rys. 1 b). Zwoje w liczbie od 2 do 12 są spiralnie zwinięte wokół osi nie mającej połączenia ze ścianą jelita. Poszczególne zwoje łączą się ze sobą a ich zewnętrzne brzegi są połączone ze ścianą jelita.
- typ pierścieniowaty - ten typ zastawki jelita powstaje w przypadku, kiedy ilość zwojów w typie spiralnym znacznie wzrośnie (Rys. 1 c). Wówczas zanika oś zastawki jelita. Poszczególne zwoje nie tworzą spirali (nie łączą się ze sobą) ale łączą się ze ścianą jelita na całym obwodzie. Ten typ zastawki występuje u rodzajów Carcharias i Mitsukurina(w niepełnej formie, w której część zwojów jest jeszcze połączona ze sobą) oraz u rodzajów Chiloscylliumi Stegostoma.
- typ zwojowaty - ten typ zastawki jelita również powstaje z typu spiralnego, poprzez zmniejszenie liczby zwojów (Rys. 1 a). W tym przypadku pojedynczy fragment tkanki jest zwinięty w zwój, Jest on nawinięty na swoją własną oś równoległą do osi jelita. Zwój ten jest połączony ze ścianą jelita na całej swojej długości, ale tylko po jednej stronie. Ten typ zastawki występuje jedynie u przedstawicieli dwóch rodzin: żarłaczowatych - Carcharhinidae oraz głowomłotowatych - Sphyrnidae.
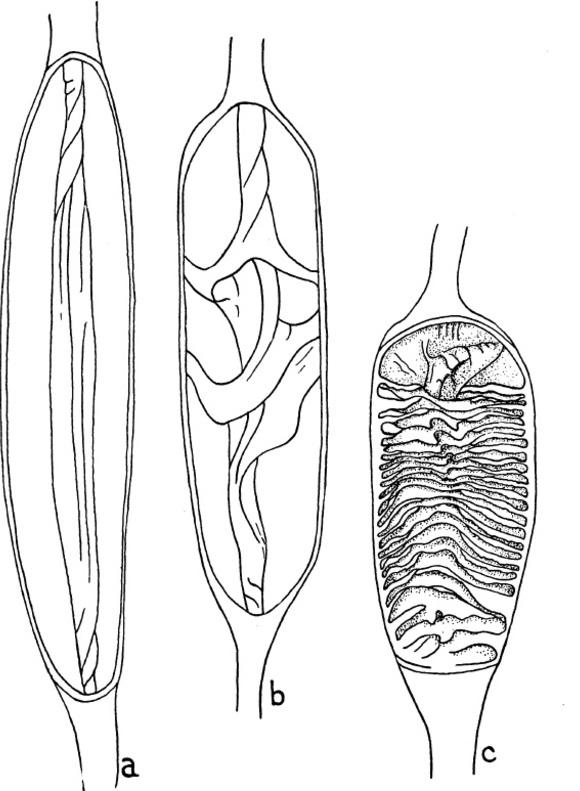 |
Rys. 1. Typy zastawek jelita występujacych u spodoustych. a - typ zwojowaty u Carcharhinus plumbeus; b - typ spiralny u Triakis scyllium; c- typ pierścieniowaty u Chiloscyllium griseum (wg White 1937).
Układ krwionośny i serce...
Podobnie jak u ryb kostnoszkieletowych u kład krwionośny u rekinów posiada tylko jeden obieg i jest typu zamkniętego.
Serce
Serce u spodoustych jest typu żylnego (płynie w nim tylko krew odtleniona). Składa się z czterech pęcherzyków:
zatoka żylna znajduje się w tylnej części serca. Przejmuje krew odtlenioną z całego organizmu transportowaną z dwóch stron przez przewody Cuviera oraz od tyłu przez żyłę wątrobową.przedsionek - w tylnej części łączy się z zatoką żylną. Jednoczenie spoczywa na komorze. Połączony jest z nią lejkowatym otworem.komora, która posiada grube ściany. Jej skurcze są główna siłą tłoczącą krew do narządów oddechowych i całego organizmu.stożek tętniczy, który przechodzi do pnia tętniczego. Skurcz stożka tętniczego zamyka zastawki prowadzące do komory uniemożliwiając cofanie się krwi z powrotem do komory. |
Serce umieszczone jest w worku osierdziowym przed wątrobą. Między nimi znajduje się przegroda poprzeczna.
Serce jest zbudowane z mięśnia poprzecznie prążkowanego sercowego, a pień tętniczy zbudowany jest z mięśni gładkich.
Pień tętniczy przechodzi w parzyste tętnice skrzelowe doprowadzające, którymi krew płynie do skrzel. Następnie tętnicami skrzelowymi odprowadzającymi natleniona krew płynie do wszystkich żywych komórek organizmu.
Układ moczowo - płciowy
Nerki
U dorosłych spodoustych występuje metameryczne pranercze. Składa się z 3 części:
- przednia część pranercza całkowicie zanika na wczesnym etapie rozwoju zarodkowego u samic. Natomiast u samców przekształca się w najądrze.
- środkowa część pranercza zanika u samic. U samców przekształca się w gruczoły Leydiga, produkujące osocze spermy. Kanaliki odchodzące od gruczołu Leydiga łączą się z nasieniowodem.
- tylna część pranercza, która pełni funkcję wydalniczą. Z każdego odcinka odchodzi kanalik prowadzący do pęcherza moczowego, który prowadzi do moczowodu pierwotnego otwierającego się do kloaki.
Narządy rozrodcze
- żeńskie narządy rozrodcze składają się z gonad - jajników (produkujących żeńskie gamety czyli komórki jajowe), jajowodu, w którym komórka jajowa jest transportowana do parzystej macicy, parzystej pochwy otwierającej się do nieparzystej kloaki.
- męskie narządy rozrodcze składają się z gonad - jąder ( produkują one gamety męskie - plemniki), najądrzy oraz nasieniowodów połączonych z kanałami odchodzącymi z gruczołów Leydiga, otwierających się do kloaki.
Tkanka chrzęstna
Szkielet wewnętrzny u wszystkich chrzęstnoszkieletowców jest zbudowany z chrząstki szklistej (hyalinowej), która zachowuje się (w przeciwieństwie do wyższych kręgowców) przez cały okres rozwoju ontogenetycznego. Jednak ze względu na znaczne obciążenia szkieletu podczas szybkiego pływania lub ataku na ofiarę, chrząstka ta ulega powierzchniowemu zwapnieniu (kalcyfikacji) przez wysycenie fosforanem wapnia - hydroksyapatytem. Dzięki temu chrząstka ryb chrzęstnoszkieletowych jest tylkonieco cięższa od chrząstki nie zwapniałej, a jednocześnie ma ona lepsze własności mechaniczne upodabniające ją do tkanki kostnej. Lekkość szkieletu ma istotne znaczenie przy braku pęcherza pławnego u wszystkichrekinów i płaszczek, u których wykształciło się wiele cech mających na celu zrekompensowanie tego defektu. Wbrew nazwie w szkielecie chrzęstnoszkieletowców występuje tkanka kostna, jednak tylko w ograniczonym zakresie do trzonów kręgów i tylko u zaawansowanych ewolucyjnie form (np: rodzaj Scyliorhinus).
Płetwy
Płetwy zbudowane są z obręczy oraz połączonych z nimi elementów szkieletu płetw.
OBRĘCZE
Wyróżnia się dwa typy obręczy:
- obręcz piersiowa u rekinów zbudowana jest zczęści kruczej i łopatkowej. Rekiny nie posiadają połączenia stawowego pomiędzy obręczą piersiową a kręgosłupem. Natomiast płaszczki posiadają lepiej rozwiniętą niż u rekinów część łopatkową połączoną stawowo z kręgosłupem.
- obręcz brzuszna również nie ma połączenia z kręgosłupem. Zbudowana jest z chrząstki łonowej i kulszowejU płaszczek z rzędu orleniokształtnych - Myliobatiformes obręcz brzuszna całkowiciezanikła. Brak u nich płetwy brzusznej, z wyjątkiem osobników męskich posiadających mixopterygia.
UWAGI:
U niektórych gatunków rekinów, obręcze brzuszne uległy zrośnięciu tworząc pomost łonowo-kulszowy, a obręcze piersiowe zrastają się ze sobą tworząc pomost łopatkowo-kruczy. Dzięki temu płetwy piersiowe mają ograniczoną ruchomość i pełnią funkcje analogiczną do skrzydeł szybowca. Podczas ruchu do przodu tworząsiłę nośną uniemożliwiającą opadanie na dno.
SZKIELET PŁETW
Płetwy służą rekinom, tak jak innym morskim zwierzętom do lokomocji.
Podział płetw ze względu na budowę ich szkieletu
- płetwy plezodyczne - szkielet płetwy zbudowany jest z actinotrichiów (keratotrichiów) i promieni podstawowych drugiego rzędu (radialiów) lub samych radialiów. Promienie podstawowesięgają dosamej krawędzi płetwy. U niektórych kopalnych rekinów występuje plezodyczna płetwa ogonowa, w której dobrze rozwinięte łuki naczyniowekręgów ogonowych wzmacniają szkielet płetwy ogonowej. U zaawansowanych płaszczek występująwtórnie plezodyczne płetwy parzyste. Radialia są u nich silnierozgałęzione, segmentowane oraz cienkie – przypominają nieco keratotrichia i zastępują je.
- płetwy semiplezodyczne – forma pośrednia pomiędzypłetwą plezodyczną a aplezodyczną. W tym typie płetwy radialia budujące szkielet nie sięgajądo krawędzi płetwy.
- płetwy aplezodyczne - szkieletpłetwy zbudowany jest jedynie z actinotrichiów (keratorichiów). W aplezodycznej płetwie ogonowej łuki naczyniowekręgów ogonowych nie wzmacniają szkieletupłetwy ogonowej. U współczesnych rekinów występujezazwyczaj aplezodyczna płetwa ogonowa.
- płetwy archipterygialne - występują u rekinów z rzędu Xenacanthiformes. Szkielet płetwy zbudowany jest z członowanego i silnie wydłużonego wyrostka metapterygialnego (powstałego z metapterygium). Wyrostek metapterygialny przebiega wzdłuż osi płetwy – radialia odchodzą od niego po obu stronach
U spodoustych - Elasmobranchii podobnie jak u ryb kostnoszkieletowych - Osteichthyes występują dwa typy płetw tj. płetwy parzyste i płetwy nieparzyste.
Płetwy parzyste
Wyróżnia się następujące płetwy parzyste: płetwę piersiową oraz płetwę brzuszną. Jak sama nazwa wskazuje są to narządy parzyste, ale mogące wtórnie zanikać w niektórych grupach płaszczek.
Płetwy parzyste zbudowane są z następujących elementów:
- promieni podstawowych pierwszego rzędu składających się z 3 płyt chrzęstnych tj. metapterygium, propterygium i mezopterygium. W szkielecie płetw brzusznych metapterygium zanika.
- promieni podstawowych drugiego rzędu (radialiów) odchodzących od promieni podstawowych pierwszego rzędu
- actinotrichiów (keratotrichiów) – są to rogopodobne twory zestawiające się z promieniami podstawowymi drugiego rzędu.
Płetwa piersiowa
Płetwy piersiowe są umiejscowione w przedniej części ciała.
Modyfikacje pierwotnego planu budowy płetwy piersiowej
- propterygium, metapterygium oraz mezopterygium zrastają się ze sobą (np. kolczak Echinorhinus);
- na metapterygium powstaje wyrostek metapterygialny (różnie rozwinięty od szczątkowego do długiego i członowanego)
- promienie podstawowe drugiego rzędu mogą być osadzone tylko:
- na mezopterygium (np. rodzaj Aculeola);
- na metapterygium (np. rodzaj Pseudocarcharias);
- na wyrostku metapterygialnym (np. kolczak Echinorhinus);
- na metapterygium i mezopterygium (np. raszpla Squatina);
- na propterygium i metapterygium;
- propterygium u płaszczek jest silnie wydłużonei łączy się za pomocą więzadła z rostrum;
- zanik promieni podstawowych pierwszego rzędu:
- mesopterygium (np. rodzaj Centrophorus)
- propterygium (np. rogatek Heterodontus)
UWAGI:
Chociaż płetwa piersiowa u aniołokształtnych Squatiniformes jest zbliżona do tej występującej u płaszczek to nie posiada ona połączenia z mózgoczaszką.
Płetwa brzuszna
Płetwa brzuszna u licznych płaszczek jest zredukowana, a u orleniokształtnych Myliobatiformes zanika całkowicie (z wyjątkiem męskich osobników, które posiadają mixopterygia).
Mixopterygia (ang. claspers) są parzystym narządem kopulacyjnym występującym u męskich osobników uwszystkich chrzęstnoszkieletowców Chondrihthyes (a przynajmniej u współczesnych) służącym do zapłodnienia wewnętrznego. Jest to narząd analogiczny do penisa wyższych kręgowców, który powstał ze zmodyfikowanych promieni podstawowych drugiego rzędu płetw brzusznych.
Płetwy nieparzyste
Wyróżnia się następujące płetwy nieparzyste: płetwa ogonowa, płetwa odbytowa oraz płetwa grzbietowa. Zazwyczaj te płetwy występują pojedynczo, ale mogą ulec wtórnemu podwojeniu lub zanikowi w niektórych grupach.
Płetwa ogonowa
Płetwa ogonowa stanowi główną siłę napędową (z wyjątkiempłaszczek, u których tą funkcję przejęła płetwa piersiowa)
Typy płetw ogonowych:
- heteroceralna płetwa ogonowa, która występuje u większości współcześnie żyjących spodoustych, a u współczesnychryb kostnoszkieletowych występuje jedynie u jesiotrowatych - Acipenseridae. Charakteryzuje się ona asymetriązewnętrzną i wewnętrzną. Grzbietowy płat płetwy ogonowej jest większy (u niektórych gatunków znacznie większy) od brzusznego. Ponadto odcinek ogonowy kręgosłupa jest skierowanydo góry (przebiega przez płat grzbietowy)
- homocerkalna płetwa ogonowa - ten typ płetwy ogonowej powszechnie występujący u ryb kostnoszkieletowych, zaś urekinów znany jest ugrup kopalnych. Charakteryzuje się symetriązewnętrzną (płaty grzbietowy i brzuszny sąmniej więcej jednakowych rozmiarów) i asymetrią wewnętrzną (odcinek ogonowy kręgosłupa przebiega przez płat grzbietowy). U niektórych współczesnych rekinów wykształciła się płetwa niemal homocerkalna.
- dyficerkalna płetwa ogonowa - typ płetwy występujący jedynie u nielicznych rekinów kopalnych (np. u rodzaju Diplodoselache), a także u ryb dwudysznych Dipnoi, czy u latimerii Latimeria. Cechą charakterystyczną jest symetria zewnętrzna i wewnętrzna.Ogonowy odcinek kręgosłupa przebiega pomiędzy płatem grzbietowym a brzusznym, wzdłuż osi płetwy.
- hipocerkalnapłetwa ogonowa, która występuje jedynie u anioła morskiego - Squatina, a także u ichtiozaurów - Ichthyosauria. Podobnie jak w przypadku płetwy heterocerkalnej mamy tu do czynienia z asymetrią zewnętrzną i wewnętrzną z tą różnicą jednak, że w płetwie hipocerkalnej płat brzuszny jest większy od grzbietowego, a odcinek ogonowy kręgosłupa przebiega przez płat brzuszny.
UWAGI:
U płaszczek z rzędu orleniokształtnych - Myliobatiformes płetwa ogonowa całkowicie zanikła - pozostaje tylko silnie wydłużony, ruchomy ogon uzbrojony w kolec jadowy. Jednak u kopalnych orleniokształtnych występowała jeszcze niewielkich rozmiarów homocerkalna płetwa ogonowa. U części gatunków rekinów zanika dolny płat płetwy ogonowej.
Płetwa odbytowa
Płetwa odbytowa posiada podobną budowę do płetwy grzbietowej, ale nigdy nie jest wzmocniona kolcem.
Kopalne rekiny z rzędu Xenacanthiformes wykształciły podwójną płetwę odbytową i silnie wydłużoną płetwę grzbietową posiadającą połączenie stawowe z mózgoczaszką. Jest to jedna z adaptacji do życia w wodach słodkich.
Przedstawiciele rzędów: koleniokształtnych Squaliformes, piłonosokształtnych Pristiophoriformes, aniołokształtnych Squatiniformes oraz płaszczki utraciły płetwę odbytową.
Płetwa grzbietowa
Szkielet płetwy grzbietowej zbudowany jest promieni chrzęstnych (pterygioforów). Mogą one występować w dużej koncentracji, a nawet zlewać się ze sobą tworząc płyty chrzęstne. Na promieniach chrzęstnych mogą być osadzone actinotrichia. Upierwotnych rekinów płetwa grzbietowa jest dodatkowo wzmocniona przez zewnętrzny lub wewnętrzny kolec
UWAGI:
U licznych rekinów występują dwie płetwy grzbietowe. U płaszczek z rzędu orleniokształtnych Myliobatiformes płetwa grzbietowa całkowicie zanikła.
Czaszka - chondrocranium
Czaszka jest typu pierwotnego i składa się z:mózgoczaszki i trzewioczaszki
Mózgoczaszka (neurocranium)
W puszce mózgowej rekinów wyróżnia się cztery okolic:
- okolica węchowa – znajdują się tu puszki węchowe otaczające pęcherzyki węchowe mózgowia. Między nimi zlokalizowany jest wysunięty do przodu dziób - rostrum. W tej części znajduje się przyczep (w hyostylicznym zawieszeniu szczęki górnej) dla więzadła łączącego chrząstkę podniebienno-kwadratową z mózgoczaszką lub powierzchnia stawowa dla tego połączenia (w amfistylicznym zawieszeniu szczęk).
- okolica oczodołowo-skroniowa – znajdują się tu oczodoły. W tylnej części oczodołu zlokalizowany jest wyrostek zaoczodołowy, na którym znajduje się powierzchnia stawowa (u rekinów posiadających amfistyliczne zawieszenie szczęk ) dla chrząstki podniebienno-kwadratowej. Od przodu oczodół jest ograniczony przez wyrostek przedoczodołowy. W tej części czaszki znajduje się również wgłębienie dla przysadki mózgowej.
- okolica słuchowa – składa się z puszki słuchowej otaczającej pęcherzyki słuchowe w mózgowiu. Na ścianach bocznych puszki słuchowej znajdują się powierzchnie stawowe dla chrząstki gnykowo-żuchwowej.okolica potyliczna – łącząca czaszkę z kręgosłupem. Zawiera otwór wielki, przez który mózg przechodzi w rdzeń kręgowy oraz pojedynczy kłykieć potyliczny lub dwa kłykcie potyliczne łączące się z pierwszym kręgiem. Połączenie to jest nieruchome, ale u niektórych płaszczek znajduje się tu staw o niewielkiej ruchomości.
UWAGI:
Sklepienie mózgoczaszki jest z reguły niekompletne – pozostaje z przodu mózgoczaszki niezamknięta przestrzeń zwana ciemiączkiem przednim.
Rostrum - dziób
Jedną z cech różniących współczesne spodouste Elasmobranchii od większości kostnoszkieletowych Osteichthyes (z wyjątkiem jesiotrokształtnych - Acipenseriformes) jest obecność rostrum - dzioba. Ten twór stanowi przedłużenie pyska i jest połączony z mózgoczaszką. Znajdują się tu przyczepy dla mięśni poruszających szczęką górną. Wszystkie rekiny i płaszczki posiadające rostrum mają subterminalny otwór gębowy. Jednak u pierwszych rekinów rostrum nie występowało, a u wielu współcześnie żyjących gatunków zanikło na skutek ewolucji wstecznej (np. u rodzaju rogatek Heterodontus, rekina wielorybiego - Rhincodon typus, czy u aniolokształtnych - Squatiniformes oraz u licznych płaszczek).
Typy rostrum występujące u rekinów i płaszczek
- rostrum w kształcie trójnoga, które zbudowane z trzech chrząstek rostralnych. W przedniej części rostrum są one zrośnięte ze sobą. Ten typrostrum występuje jedynie u lamnokształtnych - Lamniformes oraz u żarłaczokształtnych - Carcharhiniformes.
- rostrum w kształcie rowka, które zbudowane jest z tylkojednej chrząstki rostralnej. Ten typ rostrum występuje uwiększości współczesnych spodoustych (chyba, że wtórnie zanikło).
Trzewioczaszka (splanchnocranium)
Trzewioczaszkę u rekinów i płaszczek tworzy siedem łuków trzewiowych. Dwa pierwsze łuki trzewiowe utraciły pierwotną funkcję – są to łuk szczękowy (tworzący szczęki) oraz łuk gnykowy (tworzący zawieszenie szczęk). Pozostałe łuki trzewiowe nadal pełnią swoją pierwotną funkcje (oddechową) – są one zwane łukami skrzelowymi.
Łuk skrzelowy zbudowany jest z dwóch części
- gałęzi grzbietowej tworzonej przez parzyste chrząstki: epibranchiale i pharyngobranchiale.
- gałęzi brzusznej tworzonej przez parzyste chrząstki: keratobranchiale i hypobranchiale.
Hypobranchialia jednego łuku skrzelowego łączą się z następnym łukiem skrzelowym za pomocą nieparzystej chrząstki zwanej basibranchiale.
Łuk żuchwowy i gnykowy
Łuk żuchwowy – jest to zmodyfikowany pierwszy łuk trzewiowy. Podobnie jak łuki skrzelowe składa się z dwóch części
- części grzbietowej tworzonej przez parzystą chrząstkępodniebienno-kwadratową, która powstała z połączenia epibranchiale oraz pharyngobranchiale łuku skrzelowego. Gałąź górna łuku żuchwowego pełni funkcje szczęki górnej.
- części brzusznej tworzonej przezparzystą chrząstkę żuchwową (lub chrząstkę Meckela), która powstała z keratobranchiale łuku skrzelowego. Gałąź dolna łuku żuchwowego pełni funkcje szczęki dolnej (żuchwy).
Łuk gnykowy, który powstał z drugiego łuku trzewiowego, również składa się z dwóch części:
- części grzbietowej tworzonej przez chrząstkę gnykowo-żuchwową, która powstała z epibranchiale łuku skrzelowego. Chrząstka ta pełni funkcje zawieszenia szczęk i łączy łuk szczękowy z mózgoczaszką.
- części brzusznej tworzonej przez chrząstkę gnykową, która zazwyczaj podpiera dno jamy gębowej. Główny człon tej chrząstki – keratohyale powstał z keratobranchiale łuku skrzelowego.
Szczelina skrzelowa położona pomiędzy łukiem gnykowym a łukiem żuchwowym przekształciła się w tryskawkę (spiraculum). Pełni ona funkcję oddechową – umożliwia przepływ wody przez skrzela podczas spoczynku na dnie (np. podczas oczekiwania na ofiarę). Ten twór zanikł u przerazokształtnych - Chimaeriformes oraz u niektórych rekinów. Gatunki rekinów, które mają szczątkową tryskawkę lub nie mają jej w ogóle zmuszone są do nieustannego pływania lub do szukania miejsc do odpoczynku, gdzie występuje naturalny lub sztuczny nurt wody. Umożliwia im utrzymanie przepływu świeżej wody przez skrzela i podtrzymuje wymianę gazową, co zapobiega uduszeniu się.
Typy zawieszenia szczęk na mózgoczaszce:
- zawieszenie amfistyliczne (amfistylia) - ten typ zawieszenia występuje głównie u kopalnych rekinów oraz u sześcioszparokształtnych - Hexanchiformes, chlamydiokształtnych - Chlamydoselachiformes oraz rodzaju Pseudocarcharias wśród współczesnych form. Chrząstka podniebienno-kwadratowa jest połączona z mózgoczaszką w dwóch miejscach: 1. wyrostek węchowy znajdujący się na części podniebiennej chrząsrki podniebienno-kwadratowej zestawia się z okolicą węchową mózgoczaszki. 2. Processus oticus (kolejny wyrostek) znajdujący się na części kwadratowej chrząstki podniebienno-kwadratowej łączy się z wyrostkiem zaoczodołowym mózgoczaszki. Ponadto niezależnie od tych dwóch połączeń stawowych występuje jeszcze jedno połączenie z chrząstką gnykowo-żuchwową, która jest względnie mało masywna i cienka.
- zawieszenie hyostyliczne (hyostylia) - ten typ zawieszenia szczęk występuje u większości współczesnych rekinów i płaszczek oraz u ryb kostnoszkieletowych. Część podniebienna chrząstki podniebienno-kwadratowej utraciła połączenie stawowe z okolicą węchową mózgoczaszki - pozostało tylko więzadło węchowo-podniebienne.W tym przypadku chrząstka gnykowo-żuchwowa stanowi jedyne stawowe połączenie łuku żuchwowego z mózgoczaszką i jest znacznie lepiej rozwinięta niż w przypadku amfistylicznego zawieszenia.
- zawieszenie euhyostyliczne (euhyostylia) - ten typ zawieszenia szczęk występuje u płaszczek z rodziny Rhinobatidae. Łuk szczękowy jest połączony z mózgoczaszką jedynie za pomocą chrząstki gnykowo-żuchwowej. Natomiast więzadło węchowo-podniebienne całkowicie zanikło. Zapewnia to bardzo dużą ruchomość paszczy.
- zawieszenie autostyliczne (holostylia). Ten typ zawieszenia szczęk występuje u zrosłogłowych Holocephali i u ssaków. Szczęka górna jesttrwale zrośnięta z mózgoczaszką. Łuk gnykowy utracił swoją pierwotną funkcje.
Łuki skrzelowe
Pozostałe łuki trzewiowe zachowały swoją pierwotną funkcję oddechową – są to łuki skrzelowe. U rekinów i płaszczek jest zazwyczaj pięć łuków skrzelowych, ale może być także sześć łuków skrzelowych (np. sześcioszpar Hexanchus lub chlamydia Chlamydoselachus) lub nawet siedem łuków skrzelowych (np. siedmioszpar Heptranchias). Łuki skrzelowe tworzą podporę dla przegród skrzelowych. Pierwsza przegroda skrzelowa jest położona za pierwszą szczeliną skrzelową, zaś ostatnia za ostatnią.
Basibranchiale ostatniego łuku skrzelowego jest zazwyczaj większe od pozostałych i płaskie w kształcie. Osłania serce i zwane jest cardiobranchiale.
Pharyngobranciale i epibranchiale ostatniego łuku skrzelowego oraz pharyngobranchiale przedostatniego łuku skrzelowego zrastają się ze sobą tworząc tzw. oskard skrzelowy (ang. gill pickaxe). Najlepiej rozwinięty oskard skrzelowy występuje w nadrzędzie Galeoidea.
Kręgosłup i żebra
Kręgosłup u rekinów jest zbudowany z krótkich, cylindrycznych, dwuwklęsłych kręgów oraz łuków nerwowych i naczyniowych. Poszczególne kręgi połączone są wstawkami. Elementy poszczególnych łuków naczyniowych i nerwowych łączą się ze sobą tworząc pochwę otaczającą kanały naczyniowe i nerwowe. Nad łukami nerwowymi ciągnie się podłużne więzadło elastyczne. W okolicy tułowiowej kręgosłupa chrząstki łuku naczyniowego sąrozsunięte na boki tworząc wyrostki poprzeczne. Krótkie, chrzęstne żebra łączą się z nimi za pomocą więzadeł (z wyjątkiem płaszczek, u których żebra są nieruchomopołączone z wyrostkami poprzecznymi). Trzony kręgów zbudowane są z części centralnej oraz obrastającej ją w miarę rozwoju ryby – partii obwodowej. Partia obwodowa trzonu tworzy główną masę trzonu obrastając podstawy łuków naczyniowych i nerwowych oraz wstawki, w skutek czego tkwią one w trzonie.
Typy zwapnień jakim ulega trzon kręgu (patrz Rys. 1):
- astrospondylia – zwapnienia w postaci blaszek ułożonych promieniście od cylindra pierwotnego mającego tu kształt zwapniałej klepsydry.
- tektospondylia – zwapnienia mają postać koncentrycznych cylindrów ułożonych wokół cylindra pierwotnego.
- cyklospondylia – zwapnienie powierzchni trzonu w postaci prostego cylindra.
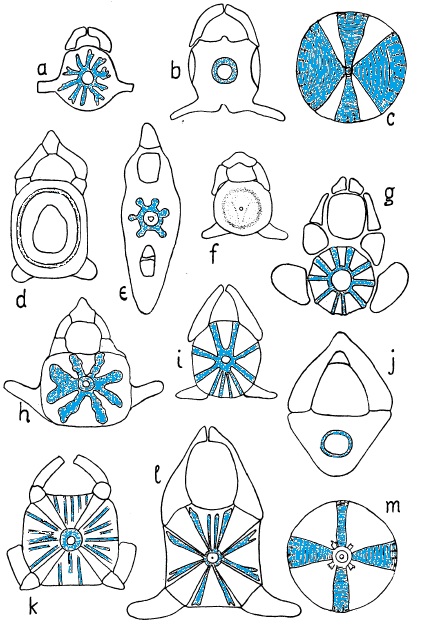 |
Rys. 1. Trzony kręgów z odcinka tułowiowego i ogonowego (tylko e). Zwapnienia zostały zaznaczone kolorem niebieskim. a - Heterodontus japonicus - tektospondylia; b - Squalus mitsukurii - cyklospondylia; c- Cetorhinus maximus - zmodyfikowana astrospondylia; d- Chimaera monstrosa - brak zwapnienia; e - Heptranchias perlo, kręg ogonowy - tektospondylia; f - Heptranchias perlo, odcinek tułowiowy - brak zwapnienia; g - Stegostoma ciliatum - astrospondylia; h - Torpedo marmoratum - tektospondylia; i - Chiloscyllium griseum - astrospondylia; j - Etmopterus lucifer - cyklospondylia; k - Carcharias taurus - astrospondylia; l - Mitsukurina owstoni - astrospondylia; m - Rhincodon typus - zmodyfikowana astrospondylia (wg White 1937)
UWAGI:
Trzon kręgu może ulegać również kostnieniu (ossyifikacji), np: urodzaju Scyliorhinus. U niektórych rekinów następuje wtórny zanik trzonów kręgów (np. chlamydia Chlamydoselachus). Zjawisko to występuje u wolno pływających rekinów, żyjących w głębinach. Z kolei u płaszczek i aniołokształtnych - Squatiniformes na pierwszych dwóch kręgach powstała chrząstka współłukowata. Zestawia ona kręgosłup z mózgoczaszką. U prymitywnych rekinów kręgosłup nie występował - funkcję kręgosłupa pełniła u nich struna grzbietowa, która w formie szczątkowej zachowała się również u współczesnych form. |
Łuski plakoidalne
U paleozoicznych rekinów występował najpierwotniejszy typ łusek – łuski lepidomorialne. Miały one prostą budowę: składały się z płytki podstawowej pokrytej zębiną i witrodentyną. W środku takiej łuski znajdowała się duża jama zębowa. Łuski tego typu miały tendencje do zrastania się u bardziej zaawansowanych form – wówczas zwane są łuskami cyklomorialnymi. Jednak ostatecznie zostały one wyparte przez łuski plakoidalne.Łuski plakoidalne pokrywają ciało spodoustych w zróżnicowanym stopniu. W niektórych grupach nastąpił wtórny zanik zębów skórnych, zwłaszcza u płaszczek.
Łuska plakoidalna składa się ze zwapniałej płytki podstawowej (plakoidalnej) tkwiącej w skórze i osadzonego na jej powierzchni ząbka. Płytka podstawowa często ulega kostnieniu. Ząbek pokryty jest jedną warstwą (prymitywne rekiny) lub kilkoma warstwami (współczesne formy) enameloidu – witrodentyny (jest to bardzo twarda tkanka zbliżona do szkliwa wyższych kręgowców), pod którą znajduje się warstwa zębiny (dentyny). W środku ząbka znajduje się jama zębowa przechodząca ku górze w kanały zębowe. Do jamy zębowej przez otwór w płytce dochodzą nerwy i naczynia krwionośne odżywiające ząbek.
M0dyfikacje zębów skórnych:
- zęby szczękowe.
- kolce płetw grzbietowych u licznych prymitywnych rekinów, obecnie głównie u rekinów z rzędu koleniokształtnych - Squaliformes i rogatkokształtnych - Heterodontiformes.
- kolce płetw piersiowych (znane tylko u paleozoicznego Doliodus problematicus).
- kolce głowowe (tylko u kopalnych np. u rodzaju Xenacanthus).
- wyrostki filtracyjne służące do odfiltrowywania planktonu z przepływającej przez aparat skrzelowy wody, u długoszpara - Cetorhinus maximus.
- krzaczkowaty twór zwieńczony zębami skórnymi u męskich osobników paleozoicznego rekina z rodzaju Stethacanthus.
- zęby rostralne (zlokalizowane na pile) u piłonosa - Pristiophorus i ryby piły - Pristis.
- płaty powiększonych łusek plakoidalnych na grzbiecie u samic płaszczek. Zapobiegają one ześlizgnięciu się samca podczas kopulacji.
Zęby szczękowe
Zęby szczękowe są zmodyfikowanymi łuskami lepidomorialnymi lub plakoidalnymi zlokalizowanymi na krawędziach szczęk. W przeciwieństwie do wyższych kręgowców nie mają one połączenia zeszkieletem szczęk, lecz są one osadzone w tkance łącznej włóknistej.
U chrzęstnoszkieletowców - Chondrichthyes występuje wymiana zębów zastępcza. Zęby ustawione są w spiralach. W każdej spirali można zaobserwować poszczególne stadia rozwojowe zęba od jego powstania do ostatniego szeregu użytkowego, znajdującego się na krawędzi szczęki. Kiedy ten ząb zostanie wyrwany np. podczas ataku na ofiarę lub wypadnie, jest on zastępowany po jakimś czasie przez ząb z drugiego rzędu. Tempo wymiany zębów jest zróżnicowane.
U licznych grup wymiana zastępcza wtórnie zanikła. Ma to zazwyczaj związek z przystosowaniami do durofagii - odżywiania się twardym pokarmem (np.: opancerzonymi organizmami z twardymi elementami szkieletu zewnętrznego).
Paleozoiczne rekiny charakteryzowały się uzębieniem homodontycznym tj. niezróżnicowanym morfologicznie ze względu na funkcje, położenie w szczęce, wiek osobnika lub histologię.
U większości współczesnych spodoustych występuje uzębienie heterodontyczne.
Typy heterodontyzmu urekinów i płaszczek:
- heterodontyzm diagonalny - zęby są zróżnicowane ze względu na umiejscowienie w szczęce tj. zęby położone w przedniej części paszczy różnią się od tych położonych w tylnej części paszczy (podobnie jak u ssaków zęby u rekinów są zestawione w serie ze względu na ich funkcje i położenie w szczęce.
- heterodontyzm dignatyczny - zęby znajdujące się w szczęce górnej różnią się od zębów położonych w tym samym miejscu w szczęce dolnej.
- kiedy uzębienie zmienia się wraz z rozwojem osobniczym tj. osobniki juwenilne (młodociane) mają inną formułę zębową lub odmienny kształt zębów niż osobniki dorosłe.u rogatka - Heterodontus
- zęby różnią się budową histologiczną: z przodu i z boku szczęk występują zęby ortodontyczne, zaś z tyłu szczęk - osteodontyczne.
UWAGI
Zęby planktonożernych rekinów charakteryzują się słabiej zaznaczonym heterodotyzmem.
Typy histologiczne zębów szczękowych::
- ortodontyczny - w tym typie zębów korona zęba jest zbudowana z ortodentyny, zaś korzeń zęba zbudowany jest z osteodentyny. W centralnej części zęba zęba znajduje się duża komora zębowa.
- osteodontyczny - korona i korzeń zęba zbudowane są tylko z osteodentyny, brak jest komory zębowej.
Podział zębów szczękowych spodoustych ze względu na kształt korony zębowej::
- zęby kladodontyczne - posiadająkilka guzków, z którychśrodkowy jest największy. Ten typ zębów jest licznie reprezentowany u rekinów (np: u paleozoicznego Cladoselache)
- zęby diplodontyczne - posiadają kilka guzków, z tą różnicą żeguzek środkowy jest najmniejszy. Ten typ zębów występuje np. u kopalnych rekinów z rodzaju Diplodoselache i u wspomnianego wcześniej Doliodus problematicus.
- zęby trójkątne - guzki boczne zanikają np: u żarłacza ludojada Carcharodon cacrcharias.
- zęby molarokształtne – będące funkcjonalnymi odpowiednikami dla trzonowców u ssaków, przypominające je kształtem.
®© GRUPA MEDIA INFORMACYJNE & ADAM NAWARA |






























